分子遺傳與創(chuàng)新中心孔凡江教授與華南農(nóng)業(yè)大學(xué)的鐘晉順教授近期在Trends in Plant Science合作系統(tǒng)地總結(jié)了禾本科和豆科復(fù)合型花序結(jié)構(gòu)遺傳調(diào)控的分子機(jī)制,,提出了有花植物進(jìn)化中復(fù)合型花序結(jié)構(gòu)調(diào)控可能存在的一般規(guī)律,。

花序結(jié)構(gòu)(inflorescence architecture)直接影響作物的產(chǎn)量(productivity)和植物的適應(yīng)度(fitness),同時(shí)也是研究植物發(fā)育和進(jìn)化研究的理想模型,。在很多有花植物中,,如擬南芥,,花序的基本組成單元是單朵花,這種簡(jiǎn)單花序可以理解為一朵朵花不斷地重復(fù)而構(gòu)成,。與之不同,,谷類和豆類作物類群植物的花序結(jié)構(gòu)為復(fù)合型,即花序的基本單元不是單朵花,,而是特化的小花序(如:禾本科小穗spikelets,;和豆科次級(jí)花序單元secondary inflorescences)。禾本科和豆科植物具有重要的農(nóng)業(yè)和生態(tài)價(jià)值,。因此,,如何理解復(fù)合型花序的建成對(duì)于生物發(fā)育和進(jìn)化研究具有重要的意義,同時(shí)也將為谷類和豆類作物的生產(chǎn)應(yīng)用提供有效的指導(dǎo)意見,。
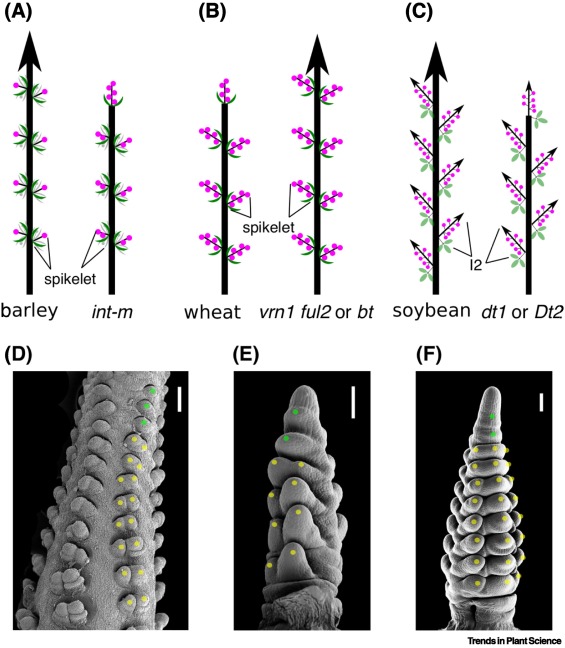
原文Fig. 1. 谷類和豆類作物形成復(fù)合型的花序,。
他們認(rèn)為特化的花序類結(jié)構(gòu)(specialized inflorescence-like structure)不能簡(jiǎn)單地解釋成分枝的形成,其本身可以作為復(fù)雜花序中的基本重復(fù)單元 – 即這種特化的花序類結(jié)構(gòu)其實(shí)可以理解為復(fù)合型的基本單元,,其自我重復(fù)與簡(jiǎn)單花序由單朵花的自我復(fù)制組成類似,。同理,這種特化的花序類結(jié)構(gòu)也可以簇生,,形成更高階的基本重復(fù)單元,,比如在玉米及其親緣類群中的小穗對(duì)(spikelet pairs)。因此,,花序類基本單元可以不斷自我重復(fù),,或者進(jìn)一步簇生形成更高階的重復(fù)單元,進(jìn)而促成花序的復(fù)雜性和導(dǎo)致復(fù)合型花序的發(fā)生,。
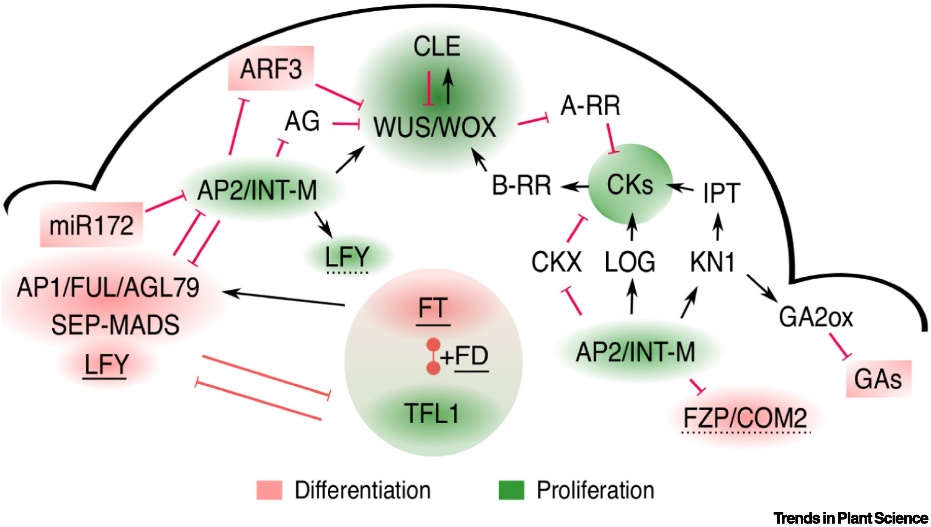
原文Fig. 2. 復(fù)合型花序頂端分生組織的調(diào)控
為了支持這個(gè)假說,,他們進(jìn)一步提出三方面的證據(jù):1. 確定性(determinate)復(fù)合型花序頂端形成一個(gè)特化花序(即禾本科小穗和豆科次級(jí)花序結(jié)構(gòu))(Fig. 1);2. 與單朵花遺傳調(diào)控類似,,花序類結(jié)構(gòu)受一系列身份基因(identity gene)的調(diào)控,,并且在禾本科和豆科中,具有高度的保守性 – 特別是TFL1-AP1/FUL/AGL79-AP2模塊具有高度保守的功能(雖然具有類群特異性,,比如利用類群特異的旁系基因paralogs)(Fig. 2),;3. 特化的小花序本身也具有一定的變異性,而這種變異的調(diào)控與花序分枝調(diào)控不同,,具有不同的機(jī)理,。同時(shí)類群間的花序基本模塊的調(diào)控機(jī)制也不盡相同,具有極大的類群特異性 (Fig. 4),。
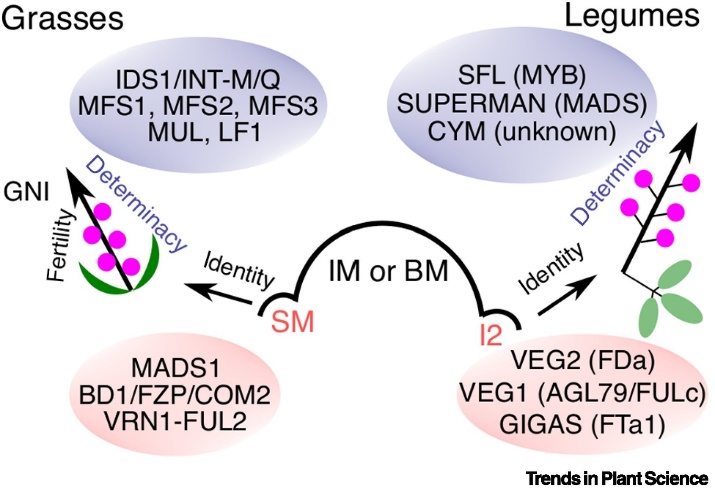
原文Fig. 4. 復(fù)合型基本單元的確定性調(diào)控
同時(shí),,該綜述還展望了禾本科和豆科花序結(jié)構(gòu)未來的研究方向和待解決的關(guān)鍵問題。我校孔凡江教授和華南農(nóng)大鐘晉順教授為論文的通訊作者,,該研究得到德國洪堡基金和國家自然科學(xué)基金的資助,。
全文鏈接:https://www.sciencedirect.com/science/article/pii/S1360138521003459?dgcid=author
 ENGLISH
ENGLISH













